瓶頸效應
瓶頸效應 (Bottleneck effect) …
遺傳漂變 (Genetic drift)
國立臺灣師範大學生命科學系研究助理林如愔
在族群中,只會有一部分的個體有機會繁衍後代,傳遞他所攜帶的基因,原因並不是他比其他個體優秀,只是純粹機率問題。這種隨機率產生的遺傳漂變是無可避免的,可以發生於所有的生物族群。遺傳漂變屬演化的基本機制,與天擇、突變、遷徙並列,而遺傳漂變究竟是如何發生的呢?

圖一 遺傳漂變的發生
在小族群中,特殊等位基因的出現頻率可能偶然地發生徹底的改變,某些等位基因可能全部由少數幾個個體攜帶,如果這些個體未能成功繁衍子嗣,便使得這些等位基因意外地遺失。如圖一中意外被踩死的綠色甲蟲,使得綠色甲殼的等位基因在這個小族群出現的頻率下降(意外地),這種等位基因頻率發生隨機變化,如同頻率漂變,即稱為遺傳漂變(genetic drift)或是等位基因漂變(allelic drift)。綠色甲蟲的例子是將問題過度單純化,現實中更多等位基因頻率的改變是不會表現在生物的外觀上,所以遺傳漂變其實一直都在發生。
蛋白質異常如何導致遺傳性聽覺障礙?
國立臺灣大學生命科學系范姜文榮編譯/國立臺灣師範大學生命科學系李冠群副教授責任編輯
編譯來源:遺伝性難聴の原因メカニズムを解明
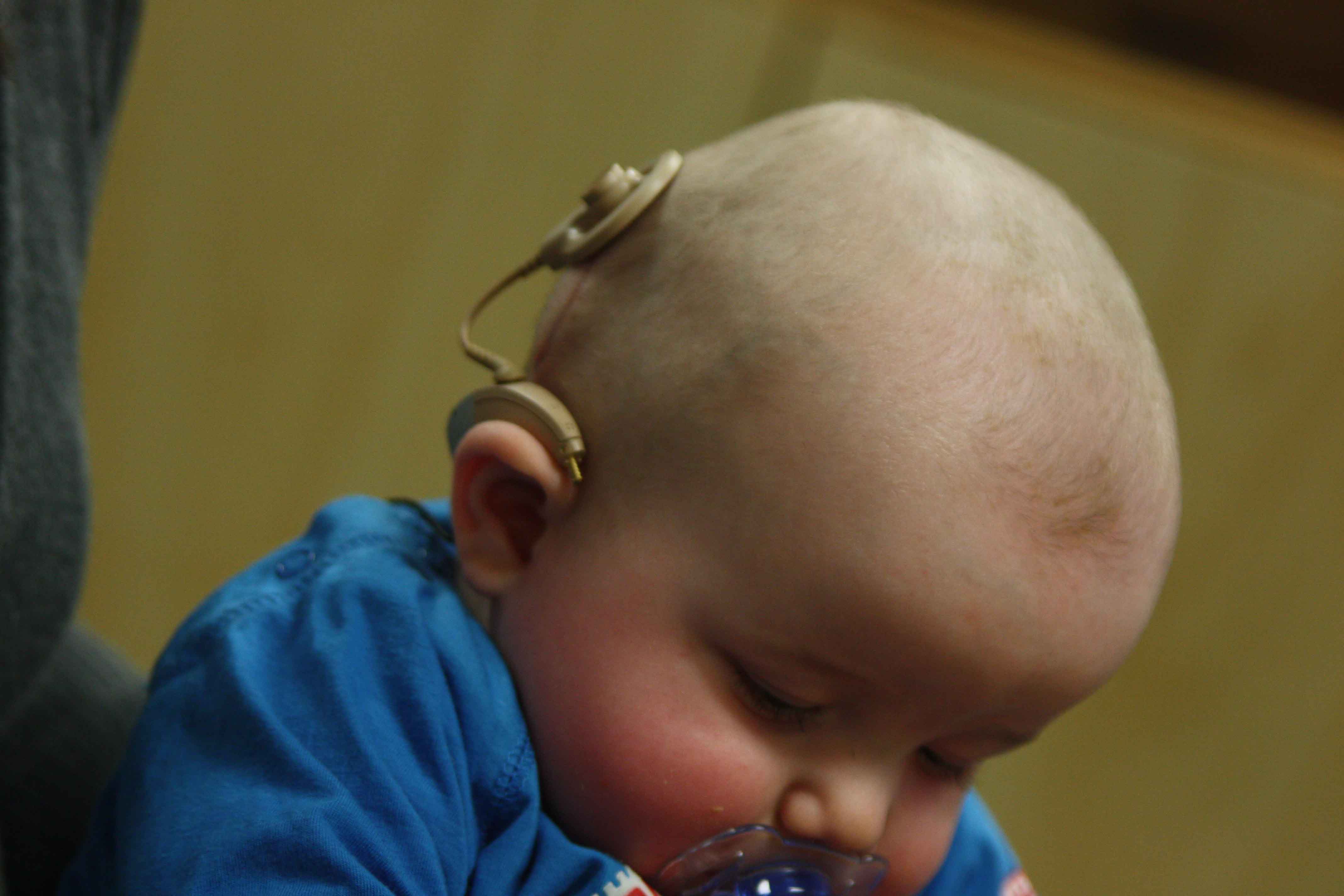
圖片來源:維基百科
聽覺障礙在新生兒的發病機率約千分之一,是先天性疾病中最常發生的疾病之一。其中半數以上都是由遺傳基因異常所造成的遺傳性聽障1,當中約50%是遺傳基因GJB2異常所導致的神經性聽障2,帶給患者語言發展或教育方面相當大的障礙。目前對此病患並無治本的治療法或治療藥。遺傳基因GJB2負責轉譯蛋白質connexin 26,它是負責內耳細胞間離子輸送的「間隙接合3」的構成要素之一,間隙接合的功能在維持內耳淋巴液的正確離子組成,聲音的震動才能轉換為神經衝動,當內耳的淋巴液組成不正確時,會導致聲音震動無法轉變成神經電訊號。但是內耳其它類別豐富的connexin蛋白質,也擔負離子輸送機能,原先認為即使蛋白質connexin 26濃度降低,輸送離子機能會得到其它類似蛋白質分子某種程度的補償,但蛋白質connexin26異常的遺傳性聽障患者仍顯示出嚴重的聽覺障礙,其原因不明。
日本順天堂大學醫學系及理化學研究所等研究團隊,為了解析蛋白質connexin 26異常如何導致聽覺障礙,製作內耳蛋白質connexin 26基因缺損之疾病模式老鼠。蛋白質connexin26異常的患者,隱性遺傳型及顯性遺傳型都會出現類似病徵。經詳細分析這兩種遺傳型態的蛋白質connexin 26基因異常老鼠之共通點,發現內耳細胞間負責離子輸送的「間隙接合區塊」之蛋白質複合體分裂嚴重,其結構大小縮小至大約27%,同時其它connexin蛋白質數量也減少至大約33%。他們認為以上結果是造成內耳無法輸送離子,導致內耳的淋巴液組成異常,聲音的震動無法轉變成神經的電訊號,形成聽覺障礙。
性別基因遺傳新發現
國立臺灣大學生命科學系助教范姜文榮
編譯來源:《ほ乳類の性決定遺伝子Sryの発現制御メカニズムの解明に成功 -人間の性分化疾患の原因解明に期待》

哺乳類性別由「X」與「Y」這2條性染色體的組合所決定,但是日本京都大學與理化學研究所的共同研究團隊發現除了X、Y性染色體外,尚有對性別決定扮演關鍵角色的其它遺傳基因。(圖片來源:維基百科)
哺乳類性別由「X」與「Y」這2條性染色體的組合所決定,但是日本京都大學與理化學研究所的共同研究團隊發現除了X、Y性染色體外,尚有對性別決定扮演關鍵角色的其它遺傳基因。
過去已知動物性別的決定機制有各種不同類型,有些魚類甚至成長後出現性別轉換,但是哺乳動物的性別決定機制嚴密,細胞核內若具有各一條X、Y性染色體,則為雄性(XY);若出現2條X性染色體,則為雌性(XX)。
雌雄的性別分化是動物為了繁衍後代的重要細胞分化過程。人類Y性染色體上遺傳基因SRY1,在雌雄未分化前之胎兒期出現短暫表現,促使XY胎兒分化為雄性,相對地, XX胎兒無SRY基因表現,則分化為雌性。但Y性染色體上的人類SRY基因或老鼠Sry基因,究竟是透過何種機制進行基因表現而促使性分化,則仍然不清楚。
劍橋參考序列 (Cambridge Reference Sequence,CRS)
國立臺灣大學醫學系呂明軒、臺北市立建國高中劉翠華教師
劍橋參考序列 (Cambridge Reference Sequence,CRS)是最廣為採用的人類粒線體 DNA 序列參考標準。
英國劍橋大學桑格博士 (Dr. Fred Sanger)是生物大分子研究領域的巨擘,發明了蛋白質與核酸的定序方法,於 1958 與 1980 年兩度獲得諾貝爾化學獎的殊榮。桑格博士的團隊於 1981 年首次發表人類粒線體 DNA (mt DNA) 全長約 $$16569$$ 個鹼基對,此序列稱為劍橋參考序列。
主要組織相容性複合物 (Major Histocompatibility Complex,MHC)
國立臺灣大學醫學系張哲睿、臺北市立建國高中劉翠華教師
主要組織相容性複合物(major histocompatibility complex,MHC)是一種細胞表面醣蛋白複合物,人類的 MHC 醣蛋白,又稱為人類白血球抗原群(human leukocyte antigens,HLA),最初是因為研究皮膚的移植和排斥反應被發現。
人類的 MHC 蛋白可以分為兩大類:第一型 MHC 分子(class I MHC)和第二型 MHC 分子(class II MHC),前者位於個體中所有有核的細胞上,後者則只分布在抗原呈現細胞(antigen-presenting cell,APC)上,例如巨噬細胞(macrophage)、B 細胞、樹突細胞(dendritic cell)等。
同源基因(Homologous genes/Homologs)
國立臺灣師範大學生命科學系研究助理鄭杏倩
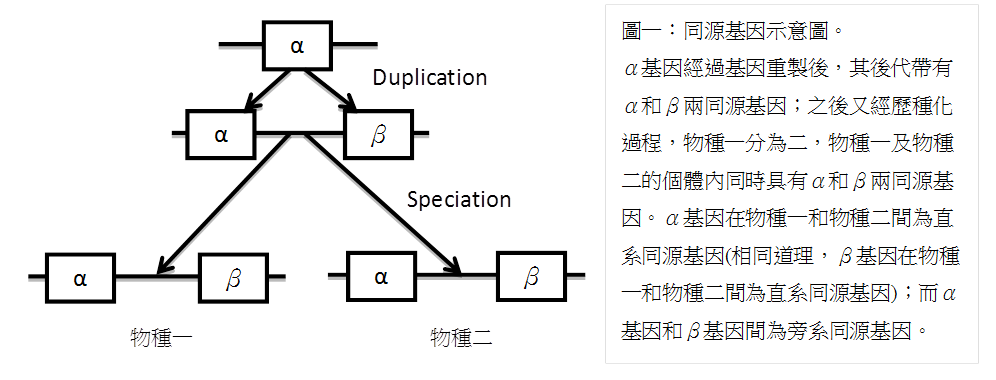
當比較兩個或多個基因時,若這些基因有共同的祖先,我們稱之為同源基因(homologous genes/homologs)。當兩同源基因存在單一物種中,也就是同源的原因是來自基因重製(gene duplication)又各自累積變異,則稱此兩基因為旁系同源基因(paralogous genes/paralogs);若兩基因存在於不同物種,也就是同源的原因來自於種化(speciation),則稱此兩基因為直系同源基因(orthologous genes/orthologs)。下圖一以簡單的示意圖闡述旁系同源與直系同源基因之定義。
原致癌基因(Proto-oncogene)
國立臺灣師範大學生命科學系研究助理胡琬琳
.jpg/800px-Cancer_cells_(1).jpg "Cancer cells") 原致癌基因(proto-oncogene)為細胞內之正常基因,其表現的蛋白質產物能促進細胞生長及分裂,當原致癌基因在諸如放射性物質、化學物質及病毒的作用影響下而突變(mutation)時,會造成其蛋白質產物過量產生或是活性增加,使細胞的生長及分裂大幅增加而導致癌症(cancer)。此時突變後的原致癌基因即稱為致癌基因(oncogene),換言之,原致癌基因是致癌基因的前身。
原致癌基因(proto-oncogene)為細胞內之正常基因,其表現的蛋白質產物能促進細胞生長及分裂,當原致癌基因在諸如放射性物質、化學物質及病毒的作用影響下而突變(mutation)時,會造成其蛋白質產物過量產生或是活性增加,使細胞的生長及分裂大幅增加而導致癌症(cancer)。此時突變後的原致癌基因即稱為致癌基因(oncogene),換言之,原致癌基因是致癌基因的前身。
原致癌基因突變為致癌基因的方式可分為三類:(1) 基因於基因組中(genome)的移動;(2) 原致癌基因的放大(amplification);(3) 原致癌基因本身或其調控區(control element)發生點突變(point mutation)。在第一類情形中,原致癌基因經轉位作用(translocation)移至不同的基因座(locus),因受到較為活化的啟動子(promoter)調控,導致其表現量增加而轉變為致癌基因;在第二類情形中,因為原致癌基因的套數(copy number)增加,導致其表現量增加而轉變為致癌基因;在第三類情形中,因為原致癌基因本身發生點突變,使其所表現的蛋白質活性增加或不易被降解(degradation),或者當原致癌基因調控區發生點突變,亦可導致其表現量增加而轉變為致癌基因。
目前研究所發現的原致癌基因大致可分為生長因子(growth factors)、受體酪氨酸激酶(receptor tyrosine kinases)、膜上非受體酪氨酸激酶(membrane mssociated non-neceptor tyrosine kinases)、G蛋白偶聯受體(G-protein coupled receptors)、絲胺酸/蘇胺酸激酶(serine/threonine kinases)、核內DNA結合/轉錄因子(nuclear DNA-binding/transcription factors)等基因。
以下分項舉例並簡介各原致癌基因:
1. 生長因子(growth factors)
SIS基因 [v-sis基因為猿猴肉瘤病毒(simian sarcoma virus)中的致癌基因] 表現血小板衍生性生長因子B鏈(PDGF B chain),此基因為最先被確認與已知細胞內基因具有同源性的致癌基因。
int-2基因表現一種與纖維母細胞生長因子相關的生長因子(FGF-related growth factor)。
KGF基因又稱HST基因,亦表現一種與纖維母細胞生長因子相關的生長因子,而且在胃癌及卡波西氏肉瘤(Kaposi’s sarcoma)細胞中被發現。
2. 受體酪氨酸激酶(receptor tyrosine kinases)
FMS基因表現血球生長激素-1 受體(colony stimulating factor-1 receptor),且被確認為一反轉錄病毒(retrovirus)的致癌基因
TRK基因表現類神經生長因子受體蛋白(NGF receptor-like protein),第一個被發現的TRK基因位於胰臟癌細胞中。
基因多效性(Gene pleiotropy)
國立臺灣師範大學生命科學系研究助理鄭杏倩
一個基因基本上控制一個性狀,然而若一個基因的突變可使多種性狀同時改變,則稱此狀況為基因多效性(gene pleiotropy),此基因則為多效性基因(pleiotropic gene)。在孟德爾的豌豆實驗中發現,具有有色種皮(seed coat)的表徵之豌豆,其花和葉腋(leaf axils)皆有色;反之,具有無色種皮表徵者,其花為白色且葉腋無色素沉澱。上述觀察用現今的知識解釋,為基因多效性的結果。種皮顏色、花瓣顏色、以及葉腋之色素沉澱三種性狀是相關聯的,控制此三種性狀之基因為同一個多效性基因。
多效性基因控制不同性狀之主要機制,是因多效性基因所生成的產物被個體內不同的細胞使用,或者,其產物具有訊息傳遞之功能並且影響許多不同的下游產物。在人類疾病中有一著名的例子:苯酮尿症(phenylketonuria;PKU),PKU是一種體染色體隱性遺傳疾病,主要是由於體內苯丙胺酸(phenylalanine)氫化(hydroxylation)成酪胺酸(tyrosine)的代謝途徑產生問題而引起的先天性代謝異常疾病。造成代謝途徑異常之基因,可同時使病患毛髮色素減少、皮膚蒼白且有智能障礙。另外,在果蠅實驗中也發現,隱性vestigial gene(vg)基因會使果蠅翅膀變短而無法飛行,也會間接改變果蠅卵巢的egg string數量、改變小盾板上的刺毛位置,並且使果蠅壽命減少。.
並不是所有的基因多效性的例子都是往相同的方向進行,有時會有拮抗的情況發生,稱為拮抗之基因多效性(antagonistic pleiotropy)。以人類為例子,p53基因之產物會使細胞停止生長,最終造成細胞死亡,因此,p53基因產物可以抑制癌細胞之生長,但也會同時抑制幹細胞(stem cell)之分裂,使人體無法自行更替老死之細胞。拮抗之基因多效性在人類老化(aging)相關的機制上,也有很多例子,G. C. Williams在1957年提出有關老化的理論,他認為某些基因產物在人體年輕的時候可以提高健康(fitness),然而當年紀漸大,此基因產物卻有相反的結果。
流行性感冒病毒的抗原漂變 (Antigenic drift)與抗原移型 (Antigenic shift)
國立臺灣師範大學生命科學系研究助理陶韻婷
流行性感冒病毒的遺傳物質為8條單股 (single strand) RNA,簡寫為ss-RNA,ss-RNA與核蛋白 (nucleoprotein, NP)相結合,纏繞成核糖核蛋白體 (ribonucleoprotein, RNP),先後由核殼蛋白層 (nucleocapsid)及基質蛋白質 (matrix protein)包覆其外,再外圍則有雙層脂膜 (lipid bilayer)圍繞,雙層脂膜又稱外套膜 (envelop),上面有兩種重要的醣蛋白,分別為血凝素蛋白 (hemagglutinin, HA)與神經胺酸酶 (neuraminidase, NA)。血凝素蛋白能與宿主細胞膜上具有唾液酸(sialic acid,又稱 N-乙醯神經胺酸 (N-acetylneuraminic acid)的醣蛋白或醣脂結合,開啟病毒進入宿主細胞的第一步。當產生的新病毒要離開宿主細胞時,神經胺酸酶會切斷新病毒與唾液酸醣蛋白的結合而釋出,再去感染其他的宿主細胞。克流感藥物能阻斷神經胺酸酶的作用,讓新合成的病毒無法釋出去感染其他正常細胞。